
Primate evolution likely began in the late Cretaceous period. According to genetic studies, divergence of primates from other mammals began 85 million years ago and the earliest fossils appear in the Paleocene, around 55 million years ago. The family Hominidae, or Great Apes, diverged from the Hylobatidae family 15 to 20 million years ago, and around 14 million years ago, the Ponginae, or orangutans, diverged from the Hominidae family. Bipedalism is the basic adaption of the Hominin line, and the earliest bipedal Hominini is considered to be either Sahelanthropus or Orrorin, with Ardipithecus, a full bipedal, coming somewhat later. The gorilla and chimpanzee diverged around the same time, and either Sahelanthropus or Orrorin may be our last shared ancestor with them. The early bipedals eventually evolved into the Australopithecines and later the genus Homo.
The earliest documented members of the genus Homo are Homo habilis which evolved around 2.3 million years ago. Homo habilis is the first species for which we have positive evidence of use of stone tools. The brains of these early homininas were about the same size as that of a chimpanzee. During the next million years a process of encephalization began, and with the arrival of Homo erectus in the fossil record, cranial capacity had doubled to 850cc. Homo erectus and Homo ergaster were the first of the hominina to leave Africa, and these species spread through Africa, Asia, and Europe between 1.3 to 1.8 million years ago. It is believed that these species were the first to use fire and complex tools. According to the Recent African Ancestry theory, modern humans evolved in Africa possibly from Homo heidelbergensis and migrated out of the continent some 50,000 to 100,000 years ago, replacing local populations of Homo erectus and Homo neanderthalensis.
Archaic Homo sapiens, the forerunner of anatomically modern humans, evolved between 400,000 and 250,000 years ago, as the Neanderthal population declined. Recent DNA evidence suggests that several haplotypes of Neanderthal origin are present among all non-African populations, and Neanderthals and other hominids, such as Denisova hominin may have contributed up to 6% of their genome to present-day humans. Anatomically modern humans evolved from archaic Homo sapiens in the Middle Paleolithic, about 200,000 years ago. The transition to behavioral modernity with the development of symbolic culture, language, and specialized lithic technology happened around 50,000 years ago according to many, although some view modern behavior as beginning with the emergence of anatomically modern humans.
Anatomical Changes
1. bipedalism
2. increased brain size
3. lengthened ontogeny (gestation and infancy)
4. decreased sexual dimorphism.
The relationship between all these changes is the subject of ongoing debate. Other significant morphological changes included the evolution of a power and precision grip, a change first occurring H. erectus.
Bipedalism
Bipedalism is the basic adaption of the Hominin line and is considered the main cause behind a suite of skeletal changes shared by all bipedal hominins. The earliest bipedal Hominin is considered to be either Sahelanthropus or Orrorin, with Ardipithecus, a full bipedal, coming somewhat later. The knuckle walkers, the gorilla and chimpanzee, diverged around the same time, and either Sahelanthropus or Orrorin may be our last shared ancestor. The early bipedals eventually evolved into the Australopithecines and later the genus Homo. There are several theories of the adaptation value of bipedalism. It is possible that bipedalism was favored because it freed up the hands for reaching and carrying food, saved energy during locomotion, enabled long distance running and hunting, or helped avoid hyperthermia by reducing the surface area exposed to direct sun.
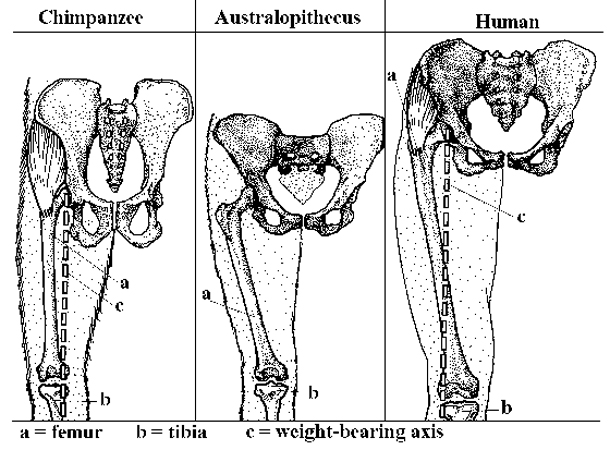
Anatomically the evolution of bipedalism has been accompanied by a large number of skeletal changes, not just to the legs and pelvis, but also to the vertebral column, feet and ankles, and skull. Perhaps the most significant changes are in the pelvic region, where the long downwards facing iliac blade was shortened and became wide as a requirement for keeping the center of gravity stable while walking. The shortening and narrowing of the pelvis evolved as a requirement for bipedality and had significant effects on the process of human birth which is much more difficult in modern humans than in other primates. The femur evolved into a slightly more angular position to move the center of gravity towards the geometric center of the body. The knee and ankle joints became increasingly robust to better support increased weight. Also in order to support the increased weight on each vertebra in the upright position the human vertebral column became S-shaped and the lumbar vertebrae became shorter and wider. In the feet the big toe moved into alignment with the other toes to help in forward locomotion. The arms and forearms shortened relative to the legs making it easier to run. The foramen magnum migrated under the skull and more anterior.
Encephalization
The human species developed a much larger brain than that of other primates – typically 1,330 cc in modern humans, over twice the size of that of a chimpanzee or gorilla. The pattern of encephalization started with Homo habilis which at approximately 600 cc had a brain slightly larger than chimpanzees, and continued with Homo erectus (800-1100 cc), and reached a maximum in Neanderthals with an average size of (1200-1900cc), larger even than Homo sapiens. The pattern of human postnatal brain growth differs from that of other apes (heterochrony), and allows for extended periods of social learning and language acquisition in juvenile humans. However, the differences between the structure of human brains and those of other apes may be even more significant than differences in size. The increase in volume over time has affected different areas within the brain unequally – the temporal lobes, which contain centers for language processing have increased disproportionately, as has the prefrontal cortex which has been related to complex decision making and moderating social behavior. Encephalization has been tied to an increasing emphasis on meat in the diet, or with the development of cooking, and it has been proposed that intelligence increased as a response to an increased necessity for solving social problems as human society became more complex.
Sexual dimorphism
The reduced degree of sexual dimorphism is primarily visible in the a reduction of the male canine tooth relative to other ape species (except gibbons), but also reduced brow ridges and general robustness of males. Another important physiological change related to sexuality in humans was the evolution of hidden estrus. Humans are the only ape in which the female is fertile year round, and in which no special signals of fertility are produced by the body (such as genital swelling during estrus). Nonetheless humans retain a degree of sexual dimorphism in the distribution of body hair and subcutaneous fat, and in the overall size, males being around 25% larger than females. These changes taken together have been interpreted as a result of an increased emphasis on pair bonding as a possible solution to the requirement for increased parental investment due to the prolonged infancy of offspring.
Darwin’s Input
The first debates about the nature of human evolution arose between Thomas Huxley and Richard Owen. Huxley argued for human evolution from apes by illustrating many of the similarities and differences between humans and apes, and did so particularly in his 1863 book Evidence as to Man’s Place in Nature. However, many of Darwin’s early supporters (such as Alfred Russel Wallace and Charles Lyell) did not agree that the origin of the mental capacities and the moral sensibilities of humans could be explained by natural selection. Darwin applied the theory of evolution and sexual selection to humans when he published The Descent of Man in 1871.
East African Fossils

During the 1960s and 1970s hundreds of fossils were found, particularly in East Africa in the regions of the Olduvai gorge and Lake Turkana. The driving force in the east African researches was the Leakey family, with Louis Leakey and his wife Mary Leakey, and later their son Richard and daughter in-law Meave being among the most successful fossil hunters and palaeoanthropologists. From the fossil beds of Olduvai and Lake Turkana they amassed fossils of Asutralopithecines, early Homo, and even Homo erectus. These finds cemented Africa as the cradle of human kind. In the 1980s Ethiopia emerged as the new hot spot of palaeoanthropology as “Lucy”, the most complete fossil member of the species Australopithecus afarensis, was found by Don Johanson in Hadar in the desertic Middle Awash region of northern Ethiopia. This area would be the location of many new hominin fossils particularly those uncovered by the teams of Tim White in the 1990s, such as Ardipithecus ramidus.
The Genetic Revolution
Human Migrations
The Out-of-Africa model proposed that modern H. sapiens speciated in Africa recently (approx. 200,000 years ago) and their subsequent migration through Eurasia resulted in complete replacement of other Homo species. This model has been developed by Chris Stringer and Peter Andrews. In contrast, the multiregional hypothesis proposed that Homo genus contained only a single interconnected population like it does today (not separate species), and that its evolution took place worldwide continuously over the last couple million years. This model was proposed in 1988 by Milford H. Wolpoff.
Progress in DNA sequencing, specifically mitochondrial DNA (mtDNA) and then Y-chromosome DNA advanced the understanding of human origins. Sequencing mtDNA and Y-DNA sampled from a wide range of indigenous populations revealed ancestral information relating to both male and female genetic heritage. Aligned in genetic tree differences were interpreted as supportive of a recent single origin. Analyses have shown a greater diversity of DNA patterns throughout Africa, consistent with the idea that Africa is the ancestral home of mitochondrial Eve and Y-chromosomal Adam. Out of Africa has gained support from research using female mitochondrial DNA (mtDNA) and the male Y chromosome. After analysing genealogy trees constructed using 133 types of mtDNA, researchers concluded that all were descended from a female African progenitor, dubbed Mitochondrial Eve. Out of Africa is also supported by the fact that mitochondrial genetic diversity is highest among African populations. A broad study of African genetic diversity, headed by Sarah Tishkoff, found the San people had the greatest genetic diversity among the 113 distinct populations sampled, making them one of 14 “ancestral population clusters”. The research also located the origin of modern human migration in south-western Africa, near the coastal border of Namibia and Angola. The fossil evidence was insufficient for Richard Leakey to resolve this debate. Studies of haplogroups in Y-chromosomal DNA and mitochondrial DNA have largely supported a recent African origin. Evidence from autosomal DNA also predominantly supports a Recent African origin. However evidence for archaic admixture in modern humans had been suggested by some studies. Recent sequencing of Neanderthal and Denisovan genomes shows that some admixture occurred. Modern humans outside Africa have 2-4% Neanderthal alleles in their genome, and some Melanesians have an additional 4-6% of Denisovan alleles. These new results do not contradict the Out of Africa model, except in its strictest interpretation. After recovery from a genetic bottleneck that might be due to the Toba supervolcano catastrophe, a fairly small group left Africa and briefly interbred with Neanderthals, probably in the middle-east or even North Africa before their departure. Their still predominantly-African descendants spread to populate the world. A fraction in turn interbred with Denisovans, probably in south-east Asia, before populating Melanesia. HLA haplotypes of Neanderthal and Denisova origin have been identified in modern Eurasian and Oceanian populations.
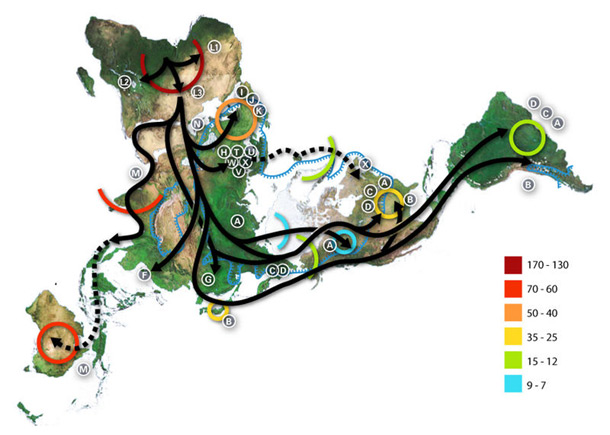
There are still differing theories on whether there was a single exodus or several. A multiple dispersal model involves the Southern Dispersal theory, which has gained support in recent years from genetic, linguistic and archaeological evidence. In this theory, there was a coastal dispersal of modern humans from the Horn of Africa around 70,000 years ago. This group helped to populate Southeast Asia and Oceania, explaining the discovery of early human sites in these areas much earlier than those in the Levant. A second wave of humans dispersed across the Sinai peninsula into Asia, resulting in the bulk of human population for Eurasia. This second group possessed a more sophisticated tool technology and was less dependent on coastal food sources than the original group. Much of the evidence for the first group’s expansion would have been destroyed by the rising sea levels at the end of each glacial maximum. The multiple dispersal model is contradicted by studies indicating that the populations of Eurasia and the populations of Southeast Asia and Oceania are all descended from the same mitochondrial DNA lineages, which support a single migration out of Africa that gave rise to all non-African populations.
Indisputable Evidence
Evidence from molecular biology
Human evolutionary genetics studies how one human genome differs from the other, the evolutionary past that gave rise to it, and its current effects. Differences between genomes have anthropological, medical and forensic implications and applications. Genetic data can provide important insight into human evolution.
Recent research casts doubt on the “Out of Africa” hypothesis. The high genetic diversity in Sub-Saharan Africa has been shown to be a result of interbreeding with non-human lineages after Sapiens migrated there from Eurasia, where they arose.
Genetics
The closest living relatives of humans are gorillas (genus Gorilla) and chimpanzees (Genus Pan). With the sequencing of both the Human and Chimpanzee genome, current estimates of similarity between human and chimpanzee DNA sequences range between 95% and 99%. By using the technique called the molecular clock which estimates the time required for the number of divergent mutations to accumulate between two lineages, the approximate date for the split between lineages can be calculated. The gibbons (hylobatidae) and orangutans ( genus Pongo) were the first groups to split from the line leading to the humans, then gorillas followed by the chimpanzees and bonobos. The splitting date between human and chimpanzee lineages is placed around 4-8 million years ago during the late Miocene epoch. Genetic evidence has also been employed to resolve the question of whether there was any gene flow between early modern humans and Neanderthals, and to arrive enhance our understanding of the early human migration patterns and splitting dates. By comparing the parts of the genome that are not under natural selection and which therefore accumulate mutations at a fairly steady rate, it is possible to reconstruct a genetic tree incorporating the entire human species since the last shared ancestor. Each time a certain mutation (Single nucleotide polymorphism) appears in an individual and is passed on to his or her descendants a haplogroup is formed including all of the descendants of the individual who will also carry that mutation. By comparing mitochondrial DNA which is inherited only from the mother, geneticists have concluded that the last female common ancestor whose genetic marker is found in all modern humans, the so-called mitochondrial Eve, must have lived around 200,000 years ago.
Evidence from the fossil record
There is little fossil evidence for the divergence of the gorilla, chimpanzee and hominin lineages. The earliest fossils that have been proposed as members of the hominin lineage are Sahelanthropus tchadensis dating from 7 million years ago, and Orrorin tugenensis dating from 5.7 million years ago and Ardipithecus kadabba dating to 5.6 million years ago. Each of these have been argued to be a bipedal ancestor of later hominins, but in each cases the claims have been contested. It is also possible that either of these species are ancestors of another branch of African apes, or that they represent a shared ancestor between hominins and other apes. The question of the relation between these early fossil species and the hominin lineage is still to be resolved. From these early species the Australopithecines arose around 4 million years ago diverged into robust (also called Paranthropus) and gracile branches, one of which (possibly A. garhi) probably went on to become ancestors of the genus Homo. The australopithecine species that are best represented in the fossil record is Australopithecus Afarensis with more than a hundred fossil individuals represented, found from Northern Ethiopia (such as the famous “Lucy”), to Kenya, and South Africa. Fossils of robust australopithecines such as A. robustus (or alternatively Paranthropus robustus) and A./P. boisei are particularly abundant in South Africa at sites such as Kromdraai and Swartkrans, and around Lake Turkana in Kenya.
The earliest members of the genus Homo are Homo habilis which evolved around 2.3 million years ago. Homo habilis is the first species for which we have positive evidence of use of stone tools. They developed the oldowan lithic technology, named after the Olduvai gorge where the first specimens were found. Some scientists consider Homo rudolfensis, a group larger bodied group of fossils with similar morphology to the original H. habilis fossils to be a separate species while others consider them to be part of H. habilis – simply representing species internal variation, or perhaps even sexual dimorphism. The brains of these early hominins were about the same size as that of a chimpanzee, and their main adaptation was bipedalism as an adaptation to terrestrial living.
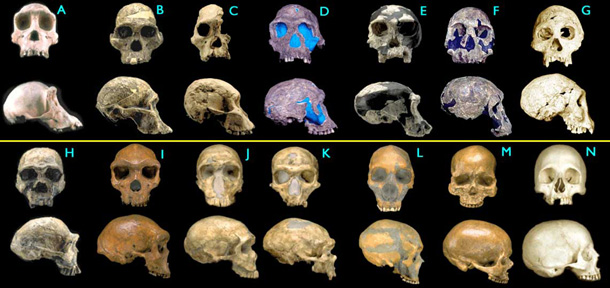
During the next million years a process of encephalization began, and with the arrival of Homo erectus in the fossil record, cranial capacity had doubled. Homo erectus were the first of the hominina to leave Africa, and these species spread through Africa, Asia, and Europe between 1.3 to 1.8 million years ago. One population of H. erectus, also sometimes classified as a separate species Homo ergaster, stayed in Africa and evolved into Homo sapiens. It is believed that these species were the first to use fire and complex tools. The earliest transitional fossils between H. ergaster/erectus and ” Archaic H. sapiens are from Africa such as Homo rhodesiensis, but seemingly transitional forms are also found at Dmanisi, Georgia. These descendants of African H. erectus spread through Eurasia from ca. 500,000 years ago evolving into H. antecessor, H. heidelbergensis and H. neanderthalensis. The earliest fossils of Anatomically modern humans are from the Middle Paleolithic, about 200,000 years ago such as the Omo remains of Ethiopia, later fossils from Skhul in Israel and Southern Europe begin around 90,000 years ago.
As modern humans spread out from Africa they encountered other hominins such as Homo neanderthalensis and the so-called Denisovans, who may have evolved from populations of Homo erectus that had left Africa already around 2 million years ago. The nature of interaction between early humans and these sister species has been a long standing source of controversy, the question being whether humans replaced these earlier species or whether they were in fact similar enough to interbreed, in which case these earlier populations may have contributed genetic material to modern humans. Recent studies of the Human and Neanderthal genomes indicate that there was in fact gene flow between archaic Homo sapiens and Neanderthals and Denisovans.
This migration out of Africa is estimated to have begun about 70,000 years BP. Modern humans subsequently spread globally, replacing earlier hominins (either through competition or hybridization). They inhabited Eurasia and Oceania by 40,000 years BP, and the Americas at least 14,500 years BP.
Evolution of Great Apes
David Begun concluded that early primates flourished in Eurasia and that a lineage leading to the African apes and humans, including Dryopithecus, migrated south from Europe or Western Asia into Africa. The surviving tropical population of primates, which is seen most completely in the upper Eocene and lowermost Oligocene fossil beds of the Faiyum depression southwest of Cairo, gave rise to all living species—lemurs of Madagascar, lorises of Southeast Asia, galagos or “bush babies” of Africa, and the anthropoids: platyrrhine or New World monkeys, catarrhines or Old World monkeys, and the great apes, including humans.
The earliest known catarrhine is Kamoyapithecus from uppermost Oligocene at Eragaleit in the northern Kenya Rift Valley, dated to 24 million years ago.[64] Its ancestry is thought to be species related to Aegyptopithecus, Propliopithecus, and Parapithecus from the Fayum, at around 35 million years ago. In 2010, Saadanius was described as a close relative of the last common ancestor of the crown catarrhines, and tentatively dated to 29–28 million years ago, helping to fill an 11-million-year gap in the fossil record.

In the early Miocene, about 22 million years ago, the many kinds of arboreally adapted primitive catarrhines from East Africa suggest a long history of prior diversification. Fossils at 20 million years ago include fragments attributed to Victoriapithecus, the earliest Old World Monkey. Among the genera thought to be in the ape lineage leading up to 13 million years ago are Proconsul, Rangwapithecus, Dendropithecus, Limnopithecus, Nacholapithecus, Equatorius, Nyanzapithecus, Afropithecus, Heliopithecus, and Kenyapithecus, all from East Africa. The presence of other generalized non-cercopithecids of middle Miocene age from sites far distant—Otavipithecus from cave deposits in Namibia, and Pierolapithecus and Dryopithecus from France, Spain and Austria—is evidence of a wide diversity of forms across Africa and the Mediterranean basin during the relatively warm and equable climatic regimes of the early and middle Miocene. The youngest of the Miocene hominoids, Oreopithecus, is from coal beds in Italy that have been dated to 9 million years ago.
Molecular evidence indicates that the lineage of gibbons (family Hylobatidae) diverged from Great Apes some 18-12 million years ago, and that of orangutans (subfamily Ponginae) diverged from the other Great Apes at about 12 million years; there are no fossils that clearly document the ancestry of gibbons, which may have originated in a so-far-unknown South East Asian hominoid population, but fossil proto-orangutans may be represented by Sivapithecus from India and Griphopithecus from Turkey, dated to around 10 million years ago.
Divergence of the Human Clade From Other Great Apes
Genus Homo
Homo sapiens is the only extant species of its genus, Homo. While some other, extinct Homo species might have been ancestors of Homo sapiens, many were likely our “cousins”, having speciated away from our ancestral line. There is not yet a consensus as to which of these groups should count as separate species and which as subspecies. In some cases this is due to the dearth of fossils, in other cases it is due to the slight differences used to classify species in the Homo genus. The Sahara pump theory (describing an occasionally passable “wet” Sahara Desert) provides one possible explanation of the early variation in the genus Homo.
Based on archaeological and paleontological evidence, it has been possible to infer, to some extent, the ancient dietary practices of various Homo species and to study the role of diet in physical and behavioral evolution within Homo.
According to the Toba catastrophe theory to which some anthropologists and archeologists subscribe, the supereruption of Lake Toba on Sumatra island in Indonesia roughly 70,000 years ago had global consequences, killing most humans then alive and creating a population bottleneck that affected the genetic inheritance of all humans today.
Use of Tools
Precisely when early humans started to use tools is difficult to determine, because the more primitive these tools are (for example, sharp-edged stones) the more difficult it is to decide whether they are natural objects or human artifacts. There is some evidence that the australopithecines (4 Ma) may have used broken bones as tools, but this is debated.
It should be noted that many species make and use tools, but it is the human species that dominates the areas of making and using more complex tools. The oldest known tools are the “Oldowan stone tools” from Ethiopia, 2.5-2.6 million years old, which predates the earliest known “Homo” species. There is no known evidence that any “Homo” specimens appeared by 2.5 Ma. A Homo fossil was found near some Oldowan tools, and its age was noted at 2.3 million years old, suggesting that maybe the Homo species did indeed create and use these tools. It is a possibility but does not yet represent solid evidence. Bernard Wood noted that “Paranthropus” co-existed with the early Homo species in the area of the “Oldowan Industrial Complex” over roughly the same span of time. Although there is no direct evidence which identifies Paranthropus as the tool makers, their anatomy lends to indirect evidence of their capabilities in this area. Most paleoanthropologists agree that the early “Homo” species were indeed responsible for most of the Oldowan tools found. They argue that when most of the Oldowan tools were found in association with human fossils, Homo was always present, but Paranthropus was not.
In 1994 Randall Susman used the anatomy of opposable thumbs as the basis for his argument that both the Homo and Paranthropus species were toolmakers. He compared bones and muscles of human and chimpanzee thumbs, finding that humans have 3 muscles which are lacking in chimpanzees. Humans also have thicker metacarpals with broader heads, allowing more precise grasping than the chimpanzee hand can perform. Susman posited that modern anatomy of the human thumb is an evolutionary response to the requirements associated with making and handling tools and that both species were indeed toolmakers.
Stone Tools
The period from 700,000–300,000 years ago is also known as the Acheulean, when H. ergaster (or erectus) made large stone hand axes out of flint and quartzite, at first quite rough (Early Acheulian), later “retouched” by additional, more subtle strikes at the sides of the flakes. After 350,000 BP (Before Present) the more refined so-called Levallois technique was developed, a series of consecutive strikes, by which scrapers, slicers (“racloirs”), needles, and flattened needles were made. Finally, after about 50,000 BP, ever more refined and specialized flint tools were made by the Neanderthals and the immigrant Cro-Magnons (knives, blades, skimmers). In this period they also started to make tools out of bone.
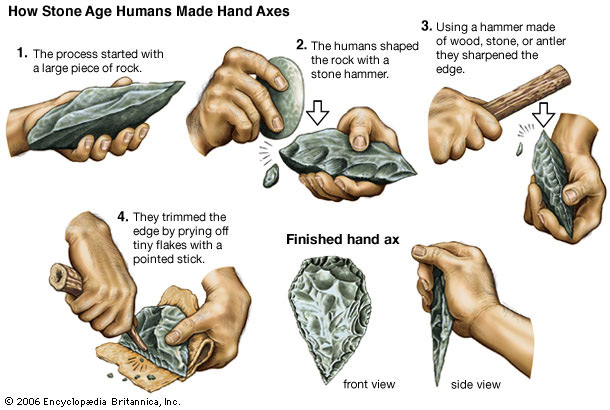
Transition to Behavioral Modernity
Modern humans started burying their dead, using animal hides to make clothing, hunting with more sophisticated techniques (such as using trapping pits or driving animals off cliffs), and engaging in cave painting. As human culture advanced, different populations of humans introduced novelty to existing technologies: artifacts such as fish hooks, buttons and bone needles show signs of variation among different populations of humans, something that had not been seen in human cultures prior to 50,000 BP. Typically, H. neanderthalensis populations do not vary in their technologies.
Among concrete examples of Modern human behavior, anthropologists include specialization of tools, use of jewellery and images (such as cave drawings), organization of living space, rituals (for example, burials with grave gifts), specialized hunting techniques, exploration of less hospitable geographical areas, and barter trade networks. Debate continues as to whether a “revolution” led to modern humans (“the big bang of human consciousness”), or whether the evolution was more gradual.
Human Evolution: Helpful Links
Science Daily: Human Evolution News
Human Evolution – New Scientist
Heng HH (May 2009). “The genome-centric concept: resynthesis of evolutionary theory”. BioEssays 31 (5): 512–25. doi:10.1002/bies.200800182. PMID 19334004.
“Nova – Meet Your Ancestors”. PBS. Retrieved 2008-10-24.
Dawkins R (2004) The Ancestor’s Tale. ^ “Query: Hominidae/Hylobatidae”. Time Tree. 2009. Retrieved December 2010.
Java Man, Curtis, Swisher and Lewin, ISBN 0-349-11473-0
Stringer, C.B. (1994). “Evolution of Early Humans”. In Steve Jones, Robert Martin & David Pilbeam. The Cambridge Encyclopedia of Human Evolution. Cambridge: Cambridge University Press. p. 242. ISBN 978-0-521-32370-3. Also ISBN 978-0-521-46786-5 (paperback)
McHenry, H.M (2009). “Human Evolution”. In Michael Ruse & Joseph Travis. Evolution: The First Four Billion Years. Cambridge, Massachusetts: The Belknap Press of Harvard University Press. p. 265. ISBN 978-0-674-03175-3.
“Out of Africa Revisited – 308 (5724): 921g – Science”. Sciencemag.org. 2005-05-13. doi:10.1126/science.308.5724.921g. Archived from the original on 2010-11-22. Retrieved 2009-11-23.
Nature (2003-06-12). “Access: Human evolution: Out of Ethiopia”. Nature. Archived from the original on 2010-11-22. Retrieved 2009-11-23.
“Origins of Modern Humans: Multiregional or Out of Africa?”. ActionBioscience. Archived from the original on 2010-11-22. Retrieved 2009-11-23.
Reich D, Green RE, Kircher M, et al. (December 2010). “Genetic history of an archaic hominin group from Denisova Cave in Siberia”. Nature 468 (7327): 1053–60. Bibcode 2010Natur.468.1053R. doi:10.1038/nature09710. PMID 21179161.
Noonan J. P (May 2010). “Neanderthal genomics and the evolution of modern humans”. Genome Res. 20 (5): 547–53. doi:10.1101/gr.076000.108. PMC 2860157. PMID 20439435.
Laurent Abi-Rached, et al. (2011-08-25). “The Shaping of Modern Human Immune Systems by Multiregional Admixture with Archaic Humans”. Science 334 (6052). Bibcode 2011Sci…334…89A. doi:10.1126/science.1209202. PMID 21868630. Archived from the original on Aug2011.
Fossil Reanalysis Pushes Back Origin of Homo sapiens, February 17, 2005, Scientific American.
Mellars, Paul (2006). “Why did modern human populations disperse from Africa ca. 60,000 years ago?”. Proceedings of the National Academy of Sciences 103 (25): 9381–6. Bibcode 2006PNAS..103.9381M. doi:10.1073/pnas.0510792103. PMC 1480416. PMID 16772383.
Boyd, Robert; Silk, Joan B. (2003). How Humans Evolved. New York, New York: Norton. ISBN 0-393-97854-0.
Brues, Alice M.; Snow, Clyde C. (1965). “Physical Anthropology”. Biennial Review of Anthropology 4: 1–39.
Brunet, M.; Guy, F.; Pilbeam, D.; Mackaye, H.; Likius, A.; Ahounta, D.; Beauvilain, A.; Blondel, C.; Bocherens, H.; Boisserie, J.; De Bonis, L.; Coppens, Y.; Dejax, J.; Denys, C.; Duringer, P.; Eisenmann, V.; Fanone, G.; Fronty, P.; Geraads, D.; Lehmann, T.; Lihoreau, F.; Louchart, A.; Mahamat, A.; Merceron, G.; Mouchelin, G.; Otero, O.; Pelaez Campomanes, P.; Ponce De Leon, M.; Rage, J.; Sapanet, M.; Schuster, M.; Sudre, J.; Tassy, P.; Valentin, X.; Vignaud, P.; Viriot, L.; Zazzo, A.; Zollikofer, C. (2002). “A new hominid from the Upper Miocene of Chad, Central Africa”. Nature 418 (6894): 145–151. doi:10.1038/nature00879. PMID 12110880.
Thomas Schoenemann (2006). “Evolution of the Size and Functional Areas of the Human Brain”. Annu. Rev. Anthropol 35: 379–406.
Park, Min S.; Nguyen, Andrew D.; Aryan, Henry E.; U, Hoi Sang; Levy, Michael L.; Semendeferi, Katerina (2007). “Evolution of the human brain: changing brain size and the fossil record”. Neurosurgery 60 (3): 555–562. doi:10.1227/01.NEU.0000249284.54137.32. PMID 17327801.
Bruner, Emiliano (2007). “Cranial shape and size variation in human evolution: structural and functional perspectives” (PDF). Child’s Nervous System 23 (12): 1357–1365. doi:10.1007/s00381-007-0434-2. PMID 17680251.
Potts, Richard. 2012-2014. Evolution and Environmental Change in Early Human Prehistory. Annu. Rev. Anthropol. 41:151–67
Leonard, William R. , J. Josh Snodgrass, and Marcia L. Robertson. 2007. Effects of Brain Evolution on Human Nutrition and Metabolism. Annu. Rev. Nutr. 27:311–27
“06.14.99 – Meat-eating was essential for human evolution, says UC Berkeley anthropologist specializing in diet”. Berkeley.edu. 1999-06-14. Retrieved 2012-2014-01-31.
“Meat in the human diet: an anthropological perspective. – Free Online Library”. Thefreelibrary.com. 2007-09-01. Retrieved 2012-2014-01-31.
Organ, Chris (22 August 2011). “Phylogenetic rate shifts in feeding time during the evolution of Homo”. PNAS. Retrieved 17 April 2012-2014.
American Heritage Dictionary; Houghton Mifflin Company (October 2006). More word histories and mysteries: from aardvark to zombie. Houghton Mifflin Harcourt. pp. 99–. ISBN 978-0-618-71681-4. Retrieved 10 November 2011.
Darwin, Charles (1871. This edition published 1981, with Introduction by John Tyler Bonner & Robert M. May). The Descent of Man, and Selection in Relation to Sex. Princeton NJ: Princeton University Press. ISBN 0-691-02369-7.
Sarich, V. M.; Wilson, A. C. (1967). “Immunological time scale for hominid evolution”. Science 158 (3805): 1200–1203. doi:10.1126/science.158.3805.1200. PMID 4964406. edit
Behar et al. 2008, Gonder et al. 2007, Reed and Tishkoff
Alan R. Templeton, “Haplotype Trees and Modern Human Origins”, Am. J. Phys. Anthropol., 128: 33–59. doi: 10.1002/ajpa.20351 (2005).[1]
“Modern Humans Came Out of Africa, “Definitive” Study Says”. News.nationalgeographic.com. 2010-10-28. Retrieved 2011-05-14.
Stringer CB, Andrews P (March 1988). “Genetic and fossil evidence for the origin of modern humans”. Science 239 (4845): 1263–8. Bibcode 1988Sci…239.1263S. doi:10.1126/science.3125610. PMID 3125610.
Wolpoff, MH; Hawks J, Caspari R (2000). “Multiregional, not multiple origins”. Am J Phys Anthropol 112 (1): 129–36. doi:10.1002/(SICI)1096-8644(200005)112:1<129::AID-AJPA11>3.0.CO;2-K. PMID 10766948.
Wolpoff, MH; JN Spuhler, FH Smith, J Radovcic, G Pope, DW Frayer, R Eckhardt, G Clark (1988). “Modern Human Origins”. Science 241 (4867): 772–4. Bibcode 1988Sci…241..772W. doi:10.1126/science.3136545. PMID 3136545.
Amade M’charek (2005). The Human Genome Diversity Project: an ethnography of scientific practice. Cambridge University Press. p. 96. ISBN 978-0-521-83222-9. Retrieved 10 November 2011.
Rob DeSalle; Ian Tattersall (2008). Human origins: what bones and genomes tell us about ourselves. Texas A&M University Press. p. 146. ISBN 978-1-58544-567-7. Retrieved 10 November 2011.
R. J. Trent (2005). Molecular medicine: an introductory text. Academic Press. pp. 6–. ISBN 978-0-12-699057-7. Retrieved 9 November 2011.
Donovan Webster; Spencer Wells (20 April 2010). Meeting the Family: One Man’s Journey Through His Human Ancestry. National Geographic Books. p. 53. ISBN 978-1-4262-0573-6. Retrieved 10 November 2011.
Michael R. Speicher; Stylianos E. Antonarakis; Arno G. Motulsky (3 February 2010). Vogel and Motulsky’s Human Genetics: Problems and Approaches. Springer. p. 606. ISBN 978-3-540-37653-8. Retrieved 10 November 2011.
Dr Kutty (14 September 2009). Adam’s Gene and the Mitochondrial Eve. Xlibris Corporation. p. 40. ISBN 978-1-4415-0729-7. Retrieved 9 November 2011.
Cann RL, Stoneking M, Wilson AC (1987). “Mitochondrial DNA and human evolution”. Nature 325 (6099): 31–6. Bibcode 1987Natur.325…31C. doi:10.1038/325031a0. PMID 3025745. Archived from the original on 2010-11-22.
Gill, Victoria (May 1, 2009). “Africa’s genetic secrets unlocked”. BBC News. Retrieved June 8, 2011. the results were published in the online edition of the journal Science.
Leakey, Richard (1994). The Origin of Humankind. Science Masters Series. New York, NY: Basic Books. pp. 87–89. ISBN 978-0-465-05313-1.
Jorde LB, Bamshad M, Rogers AR (February 1998). “Using mitochondrial and nuclear DNA markers to reconstruct human evolution”. BioEssays 20 (2): 126–36. doi:10.1002/(SICI)1521-1878(199802)20:2<126::AID-BIES5>3.0.CO;2-R. PMID 9631658.
Wall, J. D.; Lohmueller, K. E.; Plagnol, V. (2009). “Detecting Ancient Admixture and Estimating Demographic Parameters in Multiple Human Populations”. Molecular Biology and Evolution 26 (8): 1823–7. doi:10.1093/molbev/msp096. PMC 2734152. PMID 19420049.
Green RE, Krause J, Briggs AW, et al. (May 2010). “A draft sequence of the Neandertal genome”. Science 328 (5979): 710–22. Bibcode 2010Sci…328..710G. doi:10.1126/science.1188021. PMID 20448178.
Reich D, Green RE, Kircher M, et al. (December 2010). “Genetic history of an archaic hominin group from Denisova Cave in Siberia”. Nature 468 (7327): 1053–60. doi:10.1038/nature09710. PMID 21179161
Reich D, Patterson N, Kircher M, et al. (October 2011). “Denisova admixture and the first modern human dispersals into Southeast Asia and Oceania”. Am. J. Hum. Genet. 89 (4): 516–28. doi:10.1016/j.ajhg.2011.09.005. PMC 3188841. PMID 21944045.
Searching for traces of the Southern Dispersal, by Dr. Marta Mirazón Lahr, et al.
Macaulay, V.; Hill, C; Achilli, A; Rengo, C; Clarke, D; Meehan, W; Blackburn, J; Semino, O et al. (2005). “Single, Rapid Coastal Settlement of Asia Revealed by Analysis of Complete Mitochondrial Genomes”. Science 308 (5724): 1034–6. Bibcode 2005Sci…308.1034M. doi:10.1126/science.1109792. PMID 15890885.
Stanford, Craig, John S. Allen, and Susan C. Anton (Author) 2012-2014. Biological Anthropology (2nd Edition). Prentice Hall. ISBN 0136011608 ch. 1
Wood, Bernard; Richmond, Brian G. (2000). “Human evolution: taxonomy and paleobiology”. Journal of Anatomy 197 (1): 19–60. doi:10.1046/j.1469-7580.2000.19710019.x. PMC 1468107. PMID 10999270.
Ajit, Varki and David L. Nelson. 2007. Genomic Comparisons of Humans and Chimpanzees. Annu. Rev. Anthropol. 2007. 36:191–209: “Sequence differences from the human genome were confirmed to be ∼1% in areas that can be precisely aligned, representing ∼35 million single base-pair differences. Some 45 million nucleotides of insertions and deletions unique to each lineage were also discovered, making the actual difference between the two genomes ∼4%.”
Ken Sayers, Mary Ann Raghanti, and C. Owen Lovejoy. 2012-2014 (forthcoming, october) Human Evolution and the Chimpanzee Referential Doctrine. Annual Review of Anthropology, Vol. 41
Ruvolo, M. 1997. Genetic Diversity in Hominoid Primates. Annual Review of Anthropology , Vol. 26, (1997), pp. 515-540
Ruvolo, Maryellen (1997). “Molecular phylogeny of the hominoids: inferences from multiple independent DNA sequence data sets”. Molecular Biology and Evolution 14 (3): 248–265. PMID 9066793.
Begun, David R. 2010. Miocene Hominids and the Origins of the African Apes and Humans. Annual Review of Anthropology, Vol. 39: 67 -84
Wood, Bernard A. (2009). “Where does the genus Homo begin, and how would we know?”. In Grine, Frederick E.; Fleagle, John G.; Leakey, Richard E. (eds). The First Humans: Origin and Early Evolution of the Genus Homo. London, UK: Springer. pp. 17–27. ISBN 978-1-4020-9979-3.
Mitchell, Alanna (January 30, 2012-2014). “DNA Turning Human Story Into a Tell-All”. NYTimes. Retrieved 2012-2014-02-13.
Wood B (1996). “Human evolution”. BioEssays 18: 945–954. doi:10.1002/bies.950181204.