Natural selection is the gradual, non-random process by which biological traits become either more or less common in a population as a function of differential reproduction of their bearers. It is a key mechanism of evolution. The term “natural selection” was popularized by Charles Darwin who intended it to be compared with artificial selection, what we now call selective breeding.Variation exists within all populations of organisms. This occurs partly because random mutations cause changes in the genome of an individual organism, and these mutations can be passed to offspring. Throughout the individuals’ lives, their genomes interact with their environments to cause variations in traits. (The environment of a genome includes the molecular biology in the cell, other cells, other individuals, populations, species, as well as the abiotic environment.) Individuals with certain variants of the trait may survive and reproduce more than individuals with other variants. Therefore the population evolves. Factors that affect reproductive success are also important, an issue that Charles Darwin developed in his ideas on sexual selection, for example. Natural selection acts on the phenotype, or the observable characteristics of an organism, but the genetic (heritable) basis of any phenotype that gives a reproductive advantage will become more common in a population (see allele frequency). Over time, this process can result in populations that specialize for particular ecological niches and may eventually result in the emergence of new species. In other words, natural selection is an important process (though not the only process) by which evolution takes place within a population of organisms. As opposed to artificial selection, in which humans favour specific traits, in natural selection the environment acts as a sieve through which only certain variations can pass.
Natural selection is one of the cornerstones of modern biology. The term was introduced by Darwin in his influential 1859 book On the Origin of Species, in which natural selection was described as analogous to artificial selection, a process by which animals and plants with traits considered desirable by human breeders are systematically favored for reproduction. The concept of natural selection was originally developed in the absence of a valid theory of heredity; at the time of Darwin’s writing, nothing was known of modern genetics. The union of traditional Darwinian evolution with subsequent discoveries in classical and molecular genetics is termed the modern evolutionary synthesis. Natural selection remains the primary explanation for adaptive evolution.
General Principles
Natural variation occurs among the individuals of any population of organisms. Many of these differences do not affect survival (such as differences in eye color in humans), but some differences may improve the chances of survival of a particular individual. A rabbit that runs faster than others may be more likely to escape from predators, and algae that are more efficient at extracting energy from sunlight will grow faster. Something that increases an animal’s survival will often also include its reproductive rate; however, sometimes there is a trade-off between survival and current reproduction. Ultimately, what matters is total lifetime reproduction of the animal.

For example, the peppered moth exists in both light and dark colors in the United Kingdom, but during the industrial revolution many of the trees on which the moths rested became blackened by soot, giving the dark-colored moths an advantage in hiding from predators. This gave dark-colored moths a better chance of surviving to produce dark-colored offspring, and in just fifty years from the first dark moth being caught, nearly all of the moths in industrial Manchester were dark. The balance was reversed by the effect of the Clean Air Act 1956, and the dark moths became rare again, demonstrating the influence of natural selection on peppered moth evolution.If the traits that give these individuals a reproductive advantage are also heritable, that is, passed from parent to child, then there will be a slightly higher proportion of fast rabbits or efficient algae in the next generation. This is known as differential reproduction. Even if the reproductive advantage is very slight, over many generations any heritable advantage will become dominant in the population. In this way the natural environment of an organism “selects” for traits that confer a reproductive advantage, causing gradual changes or evolution of life. This effect was first described and named by Charles Darwin.
The concept of natural selection predates the understanding of genetics, the mechanism of heredity for all known life forms. In modern terms, selection acts on an organism’s phenotype, or observable characteristics, but it is the organism’s genetic make-up or genotype that is inherited. The phenotype is the result of the genotype and the environment in which the organism lives (see Genotype-phenotype distinction).
This is the link between natural selection and genetics, as described in the modern evolutionary synthesis. Although a complete theory of evolution also requires an account of how genetic variation arises in the first place (such as by mutation and sexual reproduction) and includes other evolutionary mechanisms (such as genetic drift and gene flow), natural selection appears to be the most important mechanism for creating complex adaptations in nature.
Nomenclature and Usage
The term natural selection has slightly different definitions in different contexts. It is most often defined to operate on heritable traits, because these are the traits that directly participate in evolution. However, natural selection is “blind” in the sense that changes in phenotype (physical and behavioral characteristics) can give a reproductive advantage regardless of whether or not the trait is heritable (non heritable traits can be the result of environmental factors or the life experience of the organism).Following Darwin’s primary usage the term is often used to refer to both the evolutionary consequence of blind selection and to its mechanisms. It is sometimes helpful to explicitly distinguish between selection’s mechanisms and its effects; when this distinction is important, scientists define “natural selection” specifically as “those mechanisms that contribute to the selection of individuals that reproduce”, without regard to whether the basis of the selection is heritable. This is sometimes referred to as “phenotypic natural selection”.
Traits that cause greater reproductive success of an organism are said to be selected for, whereas those that reduce success are selected against. Selection for a trait may also result in the selection of other correlated traits that do not themselves directly influence reproductive advantage. This may occur as a result of pleiotropy or gene linkage.
Fitness
The concept of fitness is central to natural selection. In broad terms, individuals that are more “fit” have better potential for survival, as in the well-known phrase “survival of the fittest”. However, as with natural selection above, the precise meaning of the term is much more subtle. Modern evolutionary theory defines fitness not by how long an organism lives, but by how successful it is at reproducing. If an organism lives half as long as others of its species, but has twice as many offspring surviving to adulthood, its genes will become more common in the adult population of the next generation.Though natural selection acts on individuals, the effects of chance mean that fitness can only really be defined “on average” for the individuals within a population. The fitness of a particular genotype corresponds to the average effect on all individuals with that genotype. Very low-fitness genotypes cause their bearers to have few or no offspring on average; examples include many human genetic disorders like cystic fibrosis.
Since fitness is an averaged quantity, it is also possible that a favorable mutation arises in an individual that does not survive to adulthood for unrelated reasons. Fitness also depends crucially upon the environment. Conditions like sickle-cell anemia may have low fitness in the general human population, but because the sickle-cell trait confers immunity from malaria, it has high fitness value in populations that have high malaria infection rates.
Types of Selection
Natural selection can act on any heritable phenotypic trait, and selective pressure can be produced by any aspect of the environment, including sexual selection and competition with members of the same or other species. However, this does not imply that natural selection is always directional and results in adaptive evolution; natural selection often results in the maintenance of the status quo by eliminating less fit variants.The unit of selection can be the individual or it can be another level within the hierarchy of biological organisation, such as genes, cells, and kin groups. There is still debate about whether natural selection acts at the level of groups or species to produce adaptations that benefit a larger, non-kin group. Likewise, there is debate as to whether selection at the molecular level prior to gene mutations and fertilization of the zygote should be ascribed to conventional natural selection because traditionally natural selection is an environmental and exterior force that acts upon a phenotype typically after birth. Some science journalists distinguish gene selection from natural selection by informally referencing selection of mutations as “pre-selection.”
Selection at a different level such as the gene can result in an increase in fitness for that gene, while at the same time reducing the fitness of the individuals carrying that gene, in a process called intragenomic conflict. Overall, the combined effect of all selection pressures at various levels determines the overall fitness of an individual, and hence the outcome of natural selection.

Natural selection occurs at every life stage of an individual. An individual organism must survive until adulthood before it can reproduce, and selection of those that reach this stage is called viability selection. In many species, adults must compete with each other for mates via sexual selection, and success in this competition determines who will parent the next generation. When individuals can reproduce more than once, a longer survival in the reproductive phase increases the number of offspring, called survival selection.The fecundity of both females and males (for example, giant sperm in certain species of Drosophila) can be limited via “fecundity selection”. The viability of produced gametes can differ, while intragenomic conflicts such as meiotic drive between the haploid gametes can result in gametic or “genic selection”. Finally, the union of some combinations of eggs and sperm might be more compatible than others; this is termed compatibility selection.
Sexual Selection
It is useful to distinguish between “ecological selection” and “sexual selection”. Ecological selection covers any mechanism of selection as a result of the environment (including relatives, e.g. kin selection, competition, and infanticide), while “sexual selection” refers specifically to competition for mates.Sexual selection can be intrasexual, as in cases of competition among individuals of the same sex in a population, or intersexual, as in cases where one sex controls reproductive access by choosing among a population of available mates. Most commonly, intrasexual selection involves male–male competition and intersexual selection involves female choice of suitable males, due to the generally greater investment of resources for a female than a male in a single offspring. However, some species exhibit sex-role reversed behavior in which it is males that are most selective in mate choice; the best-known examples of this pattern occur in some fishes of the family Syngnathidae, though likely examples have also been found in amphibian and bird species.
Some features that are confined to one sex only of a particular species can be explained by selection exercised by the other sex in the choice of a mate, for example, the extravagant plumage of some male birds. Similarly, aggression between members of the same sex is sometimes associated with very distinctive features, such as the antlers of stags, which are used in combat with other stags. More generally, intrasexual selection is often associated with sexual dimorphism, including differences in body size between males and females of a species.
Examples of Natural Selection
A well-known example of natural selection in action is the development of antibiotic resistance in microorganisms. Since the discovery of penicillin in 1928, antibiotics have been used to fight bacterial diseases. Natural populations of bacteria contain, among their vast numbers of individual members, considerable variation in their genetic material, primarily as the result of mutations. When exposed to antibiotics, most bacteria die quickly, but some may have mutations that make them slightly less susceptible. If the exposure to antibiotics is short, these individuals will survive the treatment. This selective elimination of maladapted individuals from a population is natural selection.These surviving bacteria will then reproduce again, producing the next generation. Due to the elimination of the maladapted individuals in the past generation, this population contains more bacteria that have some resistance against the antibiotic. At the same time, new mutations occur, contributing new genetic variation to the existing genetic variation. Spontaneous mutations are very rare, and advantageous mutations are even rarer. However, populations of bacteria are large enough that a few individuals will have beneficial mutations. If a new mutation reduces their susceptibility to an antibiotic, these individuals are more likely to survive when next confronted with that antibiotic.
Given enough time and repeated exposure to the antibiotic, a population of antibiotic-resistant bacteria will emerge. This new changed population of antibiotic-resistant bacteria is optimally adapted to the context it evolved in. At the same time, it is not necessarily optimally adapted any more to the old antibiotic free environment. The end result of natural selection is two populations that are both optimally adapted to their specific environment, while both perform substandard in the other environment.
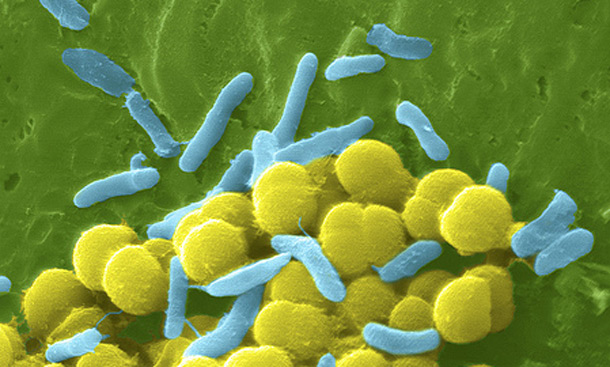
The widespread use and misuse of antibiotics has resulted in increased microbial resistance to antibiotics in clinical use, to the point that the methicillin-resistant Staphylococcus aureus (MRSA) has been described as a “superbug” because of the threat it poses to health and its relative invulnerability to existing drugs. Response strategies typically include the use of different, stronger antibiotics; however, new strains of MRSA have recently emerged that are resistant even to these drugs.This is an example of what is known as an evolutionary arms race, in which bacteria continue to develop strains that are less susceptible to antibiotics, while medical researchers continue to develop new antibiotics that can kill them. A similar situation occurs with pesticide resistance in plants and insects. Arms races are not necessarily induced by man; a well-documented example involves the spread of a gene in the butterfly Hypolimnas bolina suppressing male-killing activity by Wolbachia bacteria parasites on the island of Samoa, where the spread of the gene is known to have occurred over a period of just five years
Evolution by Means of Natural Selection
A prerequisite for natural selection to result in adaptive evolution, novel traits and speciation, is the presence of heritable genetic variation that results in fitness differences. Genetic variation is the result of mutations, recombinations and alterations in the karyotype (the number, shape, size and internal arrangement of the chromosomes). Any of these changes might have an effect that is highly advantageous or highly disadvantageous, but large effects are very rare. In the past, most changes in the genetic material were considered neutral or close to neutral because they occurred in noncoding DNA or resulted in a synonymous substitution. However, recent research suggests that many mutations in non-coding DNA do have slight deleterious effects. Although both mutation rates and average fitness effects of mutations are dependent on the organism, estimates from data in humans have found that a majority of mutations are slightly deleterious.By the definition of fitness, individuals with greater fitness are more likely to contribute offspring to the next generation, while individuals with lesser fitness are more likely to die early or fail to reproduce. As a result, alleles that on average result in greater fitness become more abundant in the next generation, while alleles that in general reduce fitness become rarer. If the selection forces remain the same for many generations, beneficial alleles become more and more abundant, until they dominate the population, while alleles with a lesser fitness disappear. In every generation, new mutations and re-combinations arise spontaneously, producing a new spectrum of phenotypes. Therefore, each new generation will be enriched by the increasing abundance of alleles that contribute to those traits that were favored by selection, enhancing these traits over successive generations.
Some mutations occur in so-called regulatory genes. Changes in these can have large effects on the phenotype of the individual because they regulate the function of many other genes. Most, but not all, mutations in regulatory genes result in non-viable zygotes. Examples of nonlethal regulatory mutations occur in HOX genes in humans, which can result in a cervical rib or polydactyly, an increase in the number of fingers or toes. When such mutations result in a higher fitness, natural selection will favor these phenotypes and the novel trait will spread in the population.
Established traits are not immutable; traits that have high fitness in one environmental context may be much less fit if environmental conditions change. In the absence of natural selection to preserve such a trait, it will become more variable and deteriorate over time, possibly resulting in a vestigial manifestation of the trait, also called evolutionary baggage. In many circumstances, the apparently vestigial structure may retain a limited functionality, or may be co-opted for other advantageous traits in a phenomenon known as preadaptation. A famous example of a vestigial structure, the eye of the blind mole rat, is believed to retain function in photoperiod perception.
Darwin’s Theory
In 1859, Charles Darwin set out his theory of evolution by natural selection as an explanation for adaptation and speciation. He defined natural selection as the “principle by which each slight variation [of a trait], if useful, is preserved”. The concept was simple but powerful: individuals best adapted to their environments are more likely to survive and reproduce. As long as there is some variation between them, there will be an inevitable selection of individuals with the most advantageous variations. If the variations are inherited, then differential reproductive success will lead to a progressive evolution of particular populations of a species, and populations that evolve to be sufficiently different eventually become different species.

Darwin’s ideas were inspired by the observations that he had made on the Beagle voyage, and by the work of a political economist, the Reverend Thomas Malthus, who in An Essay on the Principle of Population, noted that population (if unchecked) increases exponentially, whereas the food supply grows only arithmetically; thus, inevitable limitations of resources would have demographic implications, leading to a “struggle for existence”. When Darwin read Malthus in 1838 he was already primed by his work as a naturalist to appreciate the “struggle for existence” in nature and it struck him that as population outgrew resources, “favourable variations would tend to be preserved, and unfavourable ones to be destroyed. The result of this would be the formation of new species.”Here is Darwin’s own summary of the idea, which can be found in the fourth chapter of the Origin:
“If during the long course of ages and under varying conditions of life, organic beings vary at all in the several parts of their organisation, and I think this cannot be disputed; if there be, owing to the high geometrical powers of increase of each species, at some age, season, or year, a severe struggle for life, and this certainly cannot be disputed; then, considering the infinite complexity of the relations of all organic beings to each other and to their conditions of existence, causing an infinite diversity in structure, constitution, and habits, to be advantageous to them, I think it would be a most extraordinary fact if no variation ever had occurred useful to each being’s own welfare, in the same way as so many variations have occurred useful to man. But, if variations useful to any organic being do occur, assuredly individuals thus characterised will have the best chance of being preserved in the struggle for life; and from the strong principle of inheritance they will tend to produce offspring similarly characterised. This principle of preservation, I have called, for the sake of brevity, Natural Selection.”
Once he had his theory “by which to work”, Darwin was meticulous about gathering and refining evidence as his “prime hobby” before making his idea public. He was in the process of writing his “big book” to present his researches when the naturalist Alfred Russel Wallace independently conceived of the principle and described it in an essay he sent to Darwin to forward to Charles Lyell. Lyell and Joseph Dalton Hooker decided (without Wallace’s knowledge) to present his essay together with unpublished writings that Darwin had sent to fellow naturalists, and On the Tendency of Species to form Varieties; and on the Perpetuation of Varieties and Species by Natural Means of Selection was read to the Linnean Society announcing co-discovery of the principle in July 1858. Darwin published a detailed account of his evidence and conclusions in On the Origin of Species in 1859. In the 3rd edition of 1861 Darwin acknowledged that others — a notable one being William Charles Wells in 1813, and Patrick Matthew in 1831 — had proposed similar ideas, but had neither developed them nor presented them in notable scientific publications.
Darwin thought of natural selection by analogy to how farmers select crops or livestock for breeding, which he called “artificial selection”; in his early manuscripts he referred to a Nature, which would do the selection. At the time, other mechanisms of evolution such as evolution by genetic drift were not yet explicitly formulated, and Darwin believed that selection was likely only part of the story: “I am convinced that [it] has been the main, but not exclusive means of modification.” In a letter to Charles Lyell in September 1860, Darwin regretted the use of the term “Natural Selection”, preferring the term “Natural Preservation”.
For Darwin and his contemporaries, natural selection was in essence synonymous with evolution by natural selection. After the publication of On the Origin of Species, educated people generally accepted that evolution had occurred in some form. However, natural selection remained controversial as a mechanism, partly because it was perceived to be too weak to explain the range of observed characteristics of living organisms, and partly because even supporters of evolution balked at its “unguided” and non-progressive nature, a response that has been characterized as the single most significant impediment to the idea’s acceptance.
However, some thinkers enthusiastically embraced natural selection; after reading Darwin, Herbert Spencer introduced the term survival of the fittest, which became a popular summary of the theory. The fifth edition of On the Origin of Species published in 1869 included Spencer’s phrase as an alternative to natural selection, with credit given: “But the expression often used by Mr. Herbert Spencer, of the Survival of the Fittest, is more accurate, and is sometimes equally convenient.” Although the phrase is still often used by non-biologists, modern biologists avoid it because it is tautological if “fittest” is read to mean “functionally superior” and is applied to individuals rather than considered as an averaged quantity over populations.
SourcesDarwin C (1859) On the Origin of Species by Means of Natural Selection, or the Preservation of Favoured Races in the Struggle for Life John Murray, London; modern reprint Charles Darwin, Julian Huxley (2003). On The Origin of Species. Signet Classics. ISBN 0-451-52906-5. Published online at The complete work of Charles Darwin online: On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life.
Sources
Ken Miller (August, 1999). “The Peppered Moth”. Retrieved 2011-04-13.
Fisher RA (1930) The Genetical Theory of Natural Selection Clarendon Press, Oxford
Works employing or describing this usage: Endler JA (1986). Natural Selection in the Wild. Princeton, New Jersey: Princeton University Press. ISBN 0-691-00057-3.
Williams GC (1966). Adaptation and Natural Selection. Oxford University Press.
Lande R & Arnold SJ (1983) The measurement of selection on correlated characters. Evolution 37:1210-26
Futuyma DJ (2005) Evolution. Sinauer Associates, Inc., Sunderland, Massachusetts. ISBN 0-87893-187-2
Haldane, J.B.S. 1953. The measurement of natural selection. Proceedings of the 9th International Congress of Genetics. 1: 480-487
Sober E (1984; 1993) The Nature of Selection: Evolutionary Theory in Philosophical Focus University of Chicago Press ISBN 0-226-76748-5
http://www.sciencedaily.com/releases/2011/01/110125172418.htm
Modified from Christiansen FB (1984) The definition and measurement of fitness. In: Evolutionary ecology (ed. Shorrocks B) pp65–79. Blackwell Scientific, Oxford by adding survival selection in the reproductive phase
Pitnick S & Markow TA (1994) Large-male advantage associated with the costs of sperm production in Drosophila hydei, a species with giant sperm. Proc Natl Acad Sci USA 91:9277-81; Pitnick S (1996) Investment in testes and the cost of making long sperm in Drosophila. Am Nat 148:57-80
Andersson, M (1995). Sexual Selection. Princeton, New Jersey: Princeton University Press. ISBN 0-691-00057-3.
Eens, M; Pinxten, R. (2000). “Sex-role reversal in vertebrates: behavioural and endocrinological accounts”. Behav Processes 51 (1–3): 135–147. doi:10.1016/S0376-6357(00)00124-8. PMID 11074317.
Barlow, GW. (2005). “How Do We Decide that a Species is Sex-Role Reversed?”. The Quarterly Review of Biology 80 (1): 28–35. doi:10.1086/431022. PMID 15884733.
“MRSA Superbug News”. Retrieved 2006-05-06.
Schito GC (2006). “The importance of the development of antibiotic resistance in Staphylococcus aureus”. Clin Microbiol Infect 12 Suppl 1: 3–8. doi:10.1111/j.1469-0691.2006.01343.x. PMID 16445718.
Sylvain Charlat, Emily A. Hornett, James H. Fullard, Neil Davies, George K. Roderick, Nina Wedell & Gregory D. D. Hurst (2007). “Extraordinary flux in sex ratio”. Science 317 (5835): 214. doi:10.1126/science.1143369. PMID 17626876.
Kryukov, GV; Schmidt, S; Sunyaev, S (2005). “Small fitness effect of mutations in highly conserved non-coding regions”. Human Molecular Genetics 14 (15): 2221–9. doi:10.1093/hmg/ddi226. PMID 15994173.
Bejerano G, Pheasant M, Makunin I, Stephen S, Kent WJ, Mattick JS & Haussler D (2004) Ultraconserved elements in the human genome. Science 304:1321-5
Eyre-Walker, A; Woolfit, M; Phelps, T. (2006). “The distribution of fitness effects of new deleterious amino acid mutations in humans”. Genetics 173 (2): 891–900. doi:10.1534/genetics.106.057570. PMC 1526495. PMID 16547091.
Galis, F (1999). “Why do almost all mammals have seven cervical vertebrae? developmental constraints, Hox genes, and cancer”. J Exp Zool 285 (1): 19–26. doi:10.1002/(SICI)1097-010X(19990415)285:1<19::AID-JEZ3>3.0.CO;2-Z. PMID 10327647.
Zakany J, FromentalRamain C, Warot X & Duboule D (1997) Regulation of number and size of digits by posterior Hox genes: a dose-dependent mechanism with potential evolutionary implications. Proc Natl Acad Sci USA 94:13695-700
Sanyal, S; Jansen, HG; de Grip, WJ; Nevo, E; de Jong, WW. (1990). “The eye of the blind mole rat, Spalax ehrenbergi. Rudiment with hidden function?”. Invest Ophthalmol Vis Sci. 31 (7): 1398–404. PMID 2142147.
Salzburger, W; Baric, S; Sturmbauer, C. (2002). “Speciation via introgressive hybridization in East African cichlids?”. Mol Ecol 11 (3): 619–625. doi:10.1046/j.0962-1083.2001.01438.x. PMID 11918795.
Empedocles. On Nature. Book II
Lucretius. De rerum natura. Book V
Aristotle. Physics. Book II, Chapters 4 and 8
Jonathan Lear (1988). Aristotle: the desire to understand. Cambridge University Press. p. 38. ISBN 978-0-521-34762-4.
Devin Henry (2006). “Aristotle on the Mechanism of Inheritance”. Journal of the History of Biology (Springer) 39: 425–455. doi:10.1007/s10739-005-3058-y.
André Ariew (2007). “Platonic and Aristotelian Roots of Teleological Arguments in Cosmology and Biology”. In David L. Hull & Michael Ruse. The Cambridge Companion to the Philosophy of Biology. Cambridge University Press.. ISBN 978-0-521-61671-3.
Zirkle, Conway (1941). “Natural Selection before the “Origin of Species”. Proceedings of the American Philosophical Society 84 (1): 71–123.
Mehmet Bayrakdar (Third Quarter, 1983). “Al-Jahiz And the Rise of Biological Evolutionism”, The Islamic Quarterly. London.
Paul S. Agutter & Denys N. Wheatley (2008). Thinking about Life: The History and Philosophy of Biology and Other Sciences. Springer. p. 43. ISBN 1-4020-8865-5
Wallace, Alfred Russel (1870) Contributions to the Theory of Natural Selection New York: Macmillan & Co.
