
There are multiple ways to define the concept of “species”. The choice of definition is dependent on the particularities of the species concerned. For example, some species concepts apply more readily toward sexually reproducing organisms while others lend themselves better toward asexual organisms. Despite the diversity of various species concepts, these various concepts can be placed into one of three broad philosophical approaches: interbreeding, ecological and phylogenetic. The biological species concept (BSC) is a classic example of the interbreeding approach. Defined by Ernst Mayr in 1942, the BSC states that “species are groups of actually or potentially interbreeding natural populations, which are reproductively isolated from other such groups”. Despite its wide and long-term use, the BSC like others is not without controversy, for example because these concepts cannot be applied to prokaryotes, and this is called the species problem. Some researchers have attempted a unifying monistic definition of species, while others adopt a pluralistic approach and suggest that there may be a different ways to logically interpret the definition of a species.”Barriers to reproduction between two diverging sexual populations are required for the populations to become new species. Gene flow may slow this process by spreading the new genetic variants also to the other populations. Depending on how far two species have diverged since their most recent common ancestor, it may still be possible for them to produce offspring, as with horses and donkeys mating to produce mules. Such hybrids are generally infertile. In this case, closely related species may regularly interbreed, but hybrids will be selected against and the species will remain distinct. However, viable hybrids are occasionally formed and these new species can either have properties intermediate between their parent species, or possess a totally new phenotype. The importance of hybridisation in producing new species of animals is unclear, although cases have been seen in many types of animals, with the gray tree frog being a particularly well-studied example.
Speciation has been observed multiple times under both controlled laboratory conditions and in nature. In sexually reproducing organisms, speciation results from reproductive isolation followed by genealogical divergence. There are four mechanisms for speciation. The most common in animals is allopatric speciation, which occurs in populations initially isolated geographically, such as by habitat fragmentation or migration. Selection under these conditions can produce very rapid changes in the appearance and behaviour of organisms. As selection and drift act independently on populations isolated from the rest of their species, separation may eventually produce organisms that cannot interbreed.
The second mechanism of speciation is peripatric speciation, which occurs when small populations of organisms become isolated in a new environment. This differs from allopatric speciation in that the isolated populations are numerically much smaller than the parental population. Here, the founder effect causes rapid speciation after an increase in inbreeding increases selection on homozygotes, leading to rapid genetic change.
The third mechanism of speciation is parapatric speciation. This is similar to peripatric speciation in that a small population enters a new habitat, but differs in that there is no physical separation between these two populations. Instead, speciation results from the evolution of mechanisms that reduce gene flow between the two populations. Generally this occurs when there has been a drastic change in the environment within the parental species’ habitat. One example is the grass Anthoxanthum odoratum, which can undergo parapatric speciation in response to localised metal pollution from mines. Here, plants evolve that have resistance to high levels of metals in the soil. Selection against interbreeding with the metal-sensitive parental population produced a gradual change in the flowering time of the metal-resistant plants, which eventually produced complete reproductive isolation. Selection against hybrids between the two populations may cause reinforcement, which is the evolution of traits that promote mating within a species, as well as character displacement, which is when two species become more distinct in appearance.
Natural Speciation
All forms of natural speciation have taken place over the course of evolution; however it still remains a subject of debate as to the relative importance of each mechanism in driving biodiversity. One example of natural speciation is the diversity of the three-spined stickleback, a marine fish that, after the last ice age, has undergone speciation into new freshwater colonies in isolated lakes and streams. Over an estimated 10,000 generations, the sticklebacks show structural differences that are greater than those seen between different genera of fish including variations in fins, changes in the number or size of their bony plates, variable jaw structure, and color differences.
Speciation Rate
Allopatric Speciation
Allopatric speciation (from the ancient Greek allos, “other” + Greek patra, “fatherland”) or geographic speciation is speciation that occurs when biological populations of the same species become vicariant — isolated from each other to an extent that prevents or interferes with genetic interchange. This can be the result of population dispersal leading to emigration, or by geographical changes such as mountain formation, island formation, or large scale human activities (for example agricultural and civil engineering developments). The vicariant populations then undergo genotypic or phenotypic divergence as: (a) they become subjected to different selective pressures, (b) they independently undergo genetic drift, and (c) different mutations arise in the populations’ gene pools.The separate populations over time may evolve distinctly different characteristics. If the geographical barriers are later removed, members of the two populations may be unable to successfully mate with each other, at which point, the genetically isolated groups have emerged as different species. Allopatric isolation is a key factor in speciation and a common process by which new species arise. Adaptive radiation, as observed by Charles Darwin in Galapagos finches, is a consequence of allopatric speciation among island populations.
Isolating mechanisms
Allopatric speciation may occur when a species is subdivided into genetically isolated populations. Such separation commonly is referred to as vicariance. Allopatric and allopatry are terms from biogeography, referring to organisms whose ranges are entirely separate such that they do not occur in any one place together. If these organisms are closely related (e.g. sister species), such a distribution is usually the result of allopatric speciation. Separation may be attributed to either geological processes or population dispersal.
Geological processes
Geological processes can fragment a population through such events as emergence of mountain ranges, canyon formation, glacial processes, the formation or destruction of land bridges, or the subsidence of large bodies of water. On a global scale, plate tectonics are major geological factors leading to separation of populations and the resulting distribution of species.
Approximately 50,000 years ago, the Death Valley region of the western United States had a rainy climate which produced an interconnecting system of freshwater rivers and lakes. Climatic changes resulted in a drying trend that has continued for the last 10,000 years. As the lakes and rivers shrank, fish populations became geologically isolated. The few remaining (separated) springs are currently home to a variety of fish, many sharing a close common ancestor; yet each has uniquely adapted to its own particular pool.
The extent to which a geological barrier can effectively isolate a population correlates to the mobility of the organism or its offspring. For example physical barriers such as canyons may effectively block migration and dispersal of small mammals; however, have little impact on flying birds or wind-borne seeds.
Population dispersal
Population dispersal is used to describe migratory events, either in the form of range expansion (natural movement away from parents) or jump dispersal (crossing of barriers), which may lead to genetic isolation. If the smaller population fragment becomes genetically isolated from the parental group, it may be subjected to its own unique mutations, selection forces, and genetic drift effects; thus, it will follow its own evolutionary pathway. Migrations or accidental relocations (such as birds being blown off course) may lead to population fragments; whereby groups merely become separated by distance. Once gene flow between the two groups is disrupted, speciation becomes a possibility.
Examples
The African Elephant has always been regarded as a single species but, because of morphological and DNA differences, some scientists classify them into three subspecies. Researchers at the University of California, San Diego have argued that divergence due to geographical isolation has gone further, and the elephants of West Africa should be regarded as a separate species from both the savanna elephants of Central, Eastern and Southern Africa, and the forest elephants of Central Africa. A similar situation exists with the Asian Elephant, which has four distinct living sub-species.
Other cases arise where two populations that are quite distinct morphologically, and are native to different continents, have been classified as different species; but when members of one species are introduced into the other’s range, they are found to interbreed freely, showing that they were in fact only geographically isolated subspecies. This was found to be the case when the Mallard was introduced into New Zealand and interbred freely with the native Grey Duck, which had been classified as a separate species. It is controversial whether its specific status can now be retained. Mallards interbreed similarly and aggressively with the Southern African yellow-billed duck.

Observed instances
Island genetics, the tendency of small, isolated genetic pools to produce unusual traits, has been observed in many circumstances, including insular dwarfism and the radical changes among certain famous island chains, for example on Komodo. The Galápagos islands are particularly famous for their influence on Charles Darwin. During his five weeks there he heard that Galápagos tortoises could be identified by island, and noticed that Finches differed from one island to another, but it was only nine months later that he reflected that such facts could show that species were changeable. When he returned to England, his speculation on evolution deepened after experts informed him that these were separate species, not just varieties, and famously that other differing Galápagos birds were all species of finches. Though the finches were less important for Darwin, more recent research has shown the birds now known as Darwin’s finches to be a classic case of adaptive evolutionary radiation.
Peripatric Speciation
Peripatric and peripatry are terms from biogeography, referring to organisms whose ranges are closely adjacent but do not overlap, being separated where these organisms do not occur – for example a wide river or a mountain range. Such organisms are usually closely related (e.g. sister species), their distribution being the result of peripatric speciation.Peripatric speciation is a form of speciation, the formation of new species through evolution. In this form, new species are formed in isolated peripheral populations; this is similar to allopatric speciation in that populations are isolated and prevented from exchanging genes. However, peripatric speciation, unlike allopatric speciation, proposes that one of the populations is much smaller than the other. One possible consequence of peripatric speciation is that a geographically widespread ancestral species becomes paraphyletic, thereby becoming a paraspecies. The concept of a paraspecies is therefore a logical consequence of the Evolutionary Species Concept, by which one species give rise to a daughter species. The evolution of the polar bear from the brown bear is a well-documented example of a living species that gave rise to another living species through the evolution of a population located at the margin of the ancestral species’ range.
Peripatric speciation was originally proposed by Ernst Mayr, and is related to the founder effect, because small living populations may undergo selection bottlenecks. Genetic drift is often proposed to play a significant role in peripatric speciation.
Observed instances
- Mayr bird fauna
- The Australian bird Petroica multicolor
- Reproductive isolation occurs in populations of Drosophila subject to population bottlenecking
Parapatric Speciation
Parapatric speciation is a form of speciation that occurs due to apparition of dimorphism between populations of a species, and simultaneous variation in the mating habits, within a continuous geographical area. In this model, the parent species lives in a continuous habitat, in contrast with allopatric speciation & peripatric speciation where subpopulations become geographically isolated, and sympatric speciation inside the same area (and still contested).Niches in this habitat can differ along an environmental gradient, hampering gene flow, and thus creating a cline. In parapatric speciation there is no specific extrinsic barrier to gene flow. The population is continuous, but nonetheless, it does not mate randomly. Individuals are more likely to mate with their geographic neighbors than with individuals in a different part of the population’s range. In this mode, divergence may happen because of reduced gene flow within the population as a whole and varying selection pressures across the population’s range.
Examples
An example of this is the grass Anthoxanthum, which has been known to undergo parapatric speciation in such cases as mine contamination of an area. This creates a selection pressure for tolerance to those metals. Flowering time generally changes (tending toward character displacement—strong selection against interbreeding—as the hybrids are generally ill-suited to the environment) and often plants will become self-pollinating.
Similarly, a recent study provided evidence for parapatric speciation in Tennessee cave salamanders, involving divergence with gene flow between cave and surface populations.
Observed instances – ring species
- The Larus gulls form a ring species around the North Pole
- The Ensatina salamanders, which form a ring round the Central Valley in California
- The Greenish Warbler (Phylloscopus trochiloides), around the Himalayas
Sympatric Speciation
Sympatric speciation is the process through which new species evolve from a single ancestral species while inhabiting the same geographic region. In evolutionary biology and biogeography, sympatric and sympatry are terms referring to organisms whose ranges overlap or are even identical, so that they occur together at least in some places. If these organisms are closely related (e.g. sister species), such a distribution may be the result of sympatric speciation. Etymologically, sympatry is derived from the Greek roots συν (“together”, “with”) and πατρίς (“homeland” or “fatherland”). The term was invented by Poulton in 1904, who explains the derivation.Sympatric speciation is one of three traditional geographic categories for the phenomenon of speciation. Allopatric speciation is the evolution of geographically isolated populations into distinct species. In this case, divergence is facilitated by the absence of gene flow, which tends to keep populations genetically similar. Parapatric speciation is the evolution of geographically adjacent populations into distinct species. In this case, divergence occurs despite limited interbreeding where the two diverging groups come into contact. In sympatric speciation, there is no geographic constraint to interbreeding. These categories are special cases of a continuum from zero (sympatric) to complete (allopatric) spatial segregation of diverging groups.
In multicellular eukaryotic organisms, sympatric speciation is thought to be an uncommon but plausible process by which genetic divergence (through reproductive isolation) of various populations from a single parent species and inhabiting the same geographic region leads to the creation of new species.[5] In bacteria, however, the analogous process (defined as “the origin of new bacterial species that occupy definable ecological niches”) might be more common because bacteria are less constrained by the homogenizing effects of sexual reproduction and prone to comparatively dramatic and rapid genetic change through horizontal gene transfer.
Example of three-spined sticklebacks
The three-spined sticklebacks, freshwater fishes, that have been studied by Dolph Schluter (who received his Ph.D. for his work on Darwin’s finches with Peter J. Grant) and his current colleagues in British Columbia, were once thought to provide an intriguing example best explained by sympatric speciation. Schluter and colleagues found:
- Two different species of three-spined sticklebacks in each of five different lakes
- A large benthic species with a large mouth that feeds on large prey in the littoral zone
- A smaller limnetic species — with a smaller mouth — that feeds on the small plankton in open water
- DNA analysis indicates that each lake was colonized independently, presumably by a marine ancestor, after the last ice age
- DNA analysis also shows that the two species in each lake are more closely related to each other than they are to any of the species in the other lakes
- The two species in each lake are reproductively isolated; neither mates with the other.

However, aquarium tests showed:
- The benthic species from one lake will spawn with the benthic species from the other lakes and
- Likewise the limnetic species from the different lakes will spawn with each other.
- These benthic and limnetic species even display their mating preferences when presented with sticklebacks from Japanese lakes; that is, a Canadian benthic prefers a Japanese benthic over its close limnetic cousin from its own lake.
Their conclusion: in each lake, what began as a single population faced such competition for limited resources that:
- Disruptive selection — competition favoring fishes at either extreme of body size and mouth size over those nearer the mean — coupled with:
- Assortative mating — each size preferred mates like it — favored a divergence into two subpopulations exploiting different food in different parts of the lake.
The fact that this pattern of speciation occurred the same way on three separate occasions suggests strongly that ecological factors in a sympatric population can cause speciation.
However, the DNA evidence cited above is from mitochondrial DNA (mtDNA), which can often move easily between closely related species (“introgression”) when they hybridize. A more recent study, using genetic markers from the nuclear genome, shows that limnetic forms in different lakes are more closely related to each other (and to marine lineages) than to benthic forms in the same lake. The three-spine stickleback is now usually considered an example of “double invasion” (a form of allopatric speciation) in which repeated invasions of marine forms have subsequently differentiated into benthic and limnetic forms. The three-spine stickleback provides an example of how molecular biogeographic studies that rely solely on mtDNA can be misleading, and that consideration of the genealogical history of alleles from multiple unlinked markers (i.e. nuclear genes) is necessary to infer speciation histories.
Reinforcement
Reinforcement, also called the Wallace effect, is the process by which natural selection increases reproductive isolation. It may occur after two populations of the same species are separated and then come back into contact. If their reproductive isolation was complete, then they will have already developed into two separate incompatible species. If their reproductive isolation is incomplete, then further mating between the populations will produce hybrids, which may or may not be fertile. If the hybrids are infertile, or fertile but less fit than their ancestors, then there will be further reproductive isolation and speciation has essentially occurred (e.g., as in horses and donkeys.)The reasoning behind this is that if the parents of the hybrid offspring each have naturally selected traits for their own certain environments, the hybrid offspring will bear traits from both, therefore would not fit either ecological niche as well as either parent. The low fitness of the hybrids would cause selection to favor assortative mating, which would control hybridization. This is sometimes called the Wallace effect after the evolutionary biologist Alfred Russel Wallace who suggested in the late 19th century that it might be an important factor in speciation.
Conversely, if the hybrid offspring are more fit than their ancestors, then the populations will merge back into the same species within the area they are in contact.
Reinforcement favoring reproductive isolation is required for both parapatric and sympatric speciation. Without reinforcement, the geographic area of contact between different forms of the same species, called their “hybrid zone,” will not develop into a boundary between the different species. Hybrid zones are regions where diverged populations meet and interbreed. Hybrid offspring are very common in these regions, which are usually created by diverged species coming into secondary contact. Without reinforcement, the two species would have uncontrollable inbreeding. Reinforcement may be induced in artificial selection experiments as described below.
Artificial Speciation
New species have been created by domesticated animal husbandry, but the initial dates and methods of the initiation of such species are not clear. For example, domestic sheep were created by hybridisation, and no longer produce viable offspring with Ovis orientalis, one species from which they are descended. Domestic cattle, on the other hand, can be considered the same species as several varieties of wild ox, gaur, yak, etc., as they readily produce fertile offspring with them.The best-documented creations of new species in the laboratory were performed in the late 1980s. William Rice and G.W. Salt bred fruit flies, Drosophila melanogaster, using a maze with three different choices of habitat such as light/dark and wet/dry. Each generation was placed into the maze, and the groups of flies that came out of two of the eight exits were set apart to breed with each other in their respective groups. After thirty-five generations, the two groups and their offspring were isolated reproductively because of their strong habitat preferences: they mated only within the areas they preferred, and so did not mate with flies that preferred the other areas. The history of such attempts is described in Rice and Hostert (1993).
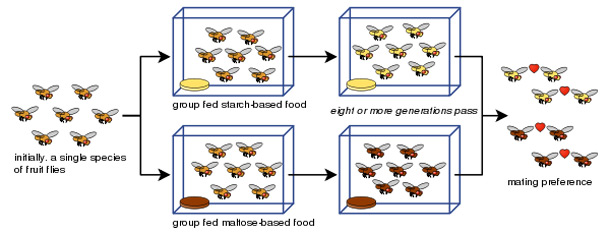
Diane Dodd was also able to show how reproductive isolation can develop from mating preferences in Drosophila pseudoobscura fruit flies after only eight generations using different food types, starch and maltose.
Hybrid Speciation
Hybridization between two different species sometimes leads to a distinct phenotype. This phenotype can also be fitter than the parental lineage and as such natural selection may then favor these individuals. Eventually, if reproductive isolation is achieved, it may lead to a separate species. However, reproductive isolation between hybrids and their parents is particularly difficult to achieve and thus hybrid speciation is considered an extremely rare event. The Mariana Mallard is known to have arisen from hybrid speciation.Hybridisation is an important means of speciation in plants, since polyploidy (having more than two copies of each chromosome) is tolerated in plants more readily than in animals. Polyploidy is important in hybrids as it allows reproduction, with the two different sets of chromosomes each being able to pair with an identical partner during meiosis. Polyploids also have more genetic diversity, which allows them to avoid inbreeding depression in small populations.
Hybridization without change in chromosome number is called homoploid hybrid speciation. It is considered very rare but has been shown in Heliconius butterflies and sunflowers. Polyploid speciation, which involves changes in chromosome number, is a more common phenomenon, especially in plant species.
Gene Transposition as a Cause
Theodosius Dobzhansky, who studied fruit flies in the early days of genetic research in 1930s, speculated that parts of chromosomes that switch from one location to another might cause a species to split into two different species. He mapped out how it might be possible for sections of chromosomes to relocate themselves in a genome. Those mobile sections can cause sterility in inter-species hybrids, which can act as a speciation pressure. In theory, his idea was sound, but scientists long debated whether it actually happened in nature. Eventually a competing theory involving the gradual accumulation of mutations was shown to occur in nature so often that geneticists largely dismissed the moving gene hypothesis.However, 2006 research shows that jumping of a gene from one chromosome to another can contribute to the birth of new species. This validates the reproductive isolation mechanism, a key component of speciation
Human Speciation
Speciation: Helpful Links
Rates of speciation in the fossil recordSpeciation rate (λ1/λ0) ratio from Bayesian Diversitree analyses.
Sources
Cook O. F. (1906). “Factors of species-formation”. Science 23 (587): 506–507. doi:10.1126/science.23.587.506. PMID 17789700.
Cook O. F. (1908). “Evolution without isolation”. American Naturalist 42: 727–731. doi:10.1086/279001.
Observed Instances of Speciation by Joseph Boxhorn. Retrieved 8 June 2009.
J.M. Baker (2005). “Adaptive speciation: The role of natural selection in mechanisms of geographic and non-geographic speciation”. Studies in History and Philosophy of Biological and Biomedical Sciences 36 (2): 303–326. doi:10.1016/j.shpsc.2005.03.005. PMID 19260194.
Kingsley, D.M. (January 2009) “From Atoms to Traits,” Scientific American, p. 57
Frank J. Sulloway (1982). “The Beagle collections of Darwin’s finches (Geospizinae)”. Bulletin of the British Museum (Natural History) Zoology Series 43 (2): 49–58. available online
Mayr, Ernst (1992). “Speciational Evolution or Puntuated Equilibrium” (PDF). Stephen jay Gould Archive. Retrieved 2012-2014-05-24.
Feder JL, Xie X, Rull J, Velez S, Forbes A, Leung B, Dambroski H, Filchak KE, Aluja M (3 May 2005). “Mayr, Dobzhansky, and Bush and the complexities of sympatric speciation in Rhagoletis”. Proc Natl Acad Sci U S A 102 (Suppl 1): 6573-6580. doi:10.1073/pnas.0502099102. PMC 1131876. PMID 15851672.
Berlocher SH, Feder JL (2002). “Sympatric speciation in phytophagous insects: moving beyond controversy?”. Annu Rev Entomol. 47: 773-815. doi:10.1146/annurev.ento.47.091201.145312. PMID 11729091.
Niemiller ML, Fitzpatrick BM, Miller BT (2008). “Recent divergence with gene flow in Tennessee cave salamanders (Plethodontidae: Gyrinophilus) inferred from gene genealogies”. Molecular Ecology 17 (9): 2258–2275. doi:10.1111/j.1365-294X.2008.03750.x. PMID 18410292.
E.B. TAYLOR, J.D. McPHAIL (2000). “Historical contingency and determinism interact to prime speciation in sticklebacks”. Proceedings of the Royal Society of London Series B 267 (1460): 2375–2384. doi:10.1098/rspb.2000.1294. PMC 1690834. PMID 11133026.[1] available online
Ramsey, J., and D. W. Schemske. 1998. Pathways, mechanisms, and rates of polyploid formation in flowering plants. Annual Review of Ecology and Systematics 29:467-501
Otto S.P., Whitton J. (2000). “Polyploidy: incidence and evolution”. Annual Review of Genetics 34: 401–437. doi:10.1146/annurev.genet.34.1.401. PMID 11092833.
Comai L (2005). “The advantages and disadvantages of being polyploid”. Nature Reviews Genetics 6 (11): 836–846. doi:10.1038/nrg1711. PMID 16304599.
Feder JL, Roethele JB, Filchak K, Niedbalski J, Romero-Severson J (1 March 2003). “Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella”. Genetics 163 (3): 939–53. PMC 1462491. PMID 12663534.
Berlocher SH, Bush GL (1982). “An electrophoretic analysis of Rhagoletis (Diptera: Tephritidae) phylogeny”. Systematic Zoology 31 (2): 136–55. doi:10.2307/2413033. JSTOR 2413033.
Ridley, M. (2003) “Speciation — What is the role of reinforcement in speciation?” adapted from Evolution 3rd edition (Boston: Blackwell Science) tutorial online
Ollerton, J. “Flowering time and the Wallace Effect” (PDF). Heredity, August 2005. Retrieved 2007-05-22.
Hiendleder S, Kaupe B, Wassmuth R, Janke A (7 May 2002). “Molecular analysis of wild and domestic sheep questions current nomenclature and provides evidence for domestication from two different subspecies”. Proc Biol Sci. 269 (1494): 893-904. doi:10.1098/rspb.2002.1975. PMC 1690972. PMID 12028771.
Nowak, R. (1999). Walker’s Mammals of the World (6th ed.). Baltimore: Johns Hopkins University Press.
Rice WR, Salt GW (1988). “Speciation via disruptive selection on habitat preference: experimental evidence”. The American Naturalist 131: 911–917. doi:10.1086/284831.
Rice WR, Hostert EE (1993). “Laboratory experiments on speciation: What have we learned in forty years?”. Evolution 47 (6): 1637–1653. doi:10.2307/2410209. JSTOR 2410209.
Gavrilets S (October 2003). “Perspective: models of speciation: what have we learned in 40 years?”. Evolution 57 (10): 2197-2215. doi:10.1554/02-727. PMID 14628909.
Dodd, D.M.B. (1989). “Reproductive isolation as a consequence of adaptive divergence in Drosophila pseudoobscura”. Evolution 43 (6): 1308–1311. doi:10.2307/2409365. JSTOR 2409365.
Kirkpatrick M, Ravigné V (2002). “Speciation by Natural and Sexual Selection: Models and Experiments”. The American Naturalist 159: S22–S35. doi:10.1086/338370. PMID 18707367.
Koukou K, Pavlikaki H, Kilias G, Werren JH, Bourtzis K, Alahiotis SN (January 2006). “Influence of antibiotic treatment and Wolbachia curing on sexual isolation among Drosophila melanogaster cage populations”. Evolution 60 (1): 87-96.
Provine WB (1 July 2004). “Ernst Mayr: Genetics and speciation”. Genetics 167 (3): 1041–6. PMC 1470966. PMID 15280221.
Templeton AR (1 April 1980). “The theory of speciation via the founder principle”. Genetics 94 (4): 1011–38. PMC 1214177. PMID 6777243
Antonovics J (July 2006). “Evolution in closely adjacent plant populations X: long-term persistence of prereproductive isolation at a mine boundary”. Heredity 97 (1): 33–7. doi:10.1038/sj.hdy.6800835. PMID 16639420.
MATTHEW L. NIEMILLER, BENJAMIN M. FITZPATRICK, BRIAN T. MILLER (2008). “Recent divergence with gene flow in Tennessee cave salamanders (Plethodontidae: Gyrinophilus) inferred from gene genealogies”. Molecular Ecology 17 (9): 2258–2275. doi:10.1111/j.1365-294X.2008.03750.x. PMID 18410292.
Parapatric speciation” in Understanding Evolution at evolution.berkeley.edu
Poulton, E. B. 1904. What is a species? Proceedings of the Entomological Society of London 1903:lxxvii-cxvi.
Futuyma, D. J. 2009. Evolution (2nd edition). Sinauer Associates, Inc.
Fitzpatrick, B. M., J. A. Fordyce, and S. Gavrilets. 2008. What, if anything, is sympatric speciation? Journal of Evolutionary Biology 21: 1452-1459.
Bolnick, D. I. and B. M. Fitzpatrick. 2007. Sympatric speciation: Models and empirical evidence. Annual Review of Ecology, Evolution and Systematics 38: 459-487.
King, Stansfield, Mulligan (2006). Dictionary of Genetics (7th ed.). Oxford University Press.
John Maynard Smith (1966). “Sympatric Speciation”. American Naturalist 100 (916): 637–650.
Thierry Lodé “La guerre des sexes chez les animaux” Eds O Jacob, Paris, 2006
*Huber, SK; De León, LF; Hendry, AP; Bermingham, E; Podos, J (2007). “Reproductive isolation of sympatric morphs in a population of Darwin’s finches”. Proc. Biol. Sci. 274 (1619): 1709–14. doi:10.1098/rspb.2007.0224. PMC 2493575. PMID 17504742
*Kingston, T.; Rossiter, S.J. (2004). “Harmonic hopping in Wallacea’s bats”. Nature 429 (6992): 654–7. doi:10.1038/nature02487. PMID 15190351
Begon, Townsend, Harper: Ecology – From individuals to ecosystems, 4th ed., p.10
Campbell, Neil; Reese, Jane (2002). Biology Sixth Edition (6 ed.). Benjamin Cummings. ISBN 0-8053-6624-5.
Fickel, J.; Lieckfeldt, D.; Ratanakorn, P.; Pitra, C. (2007). “Distribution of haplotypes and microsatellite alleles among Asian elephants (Elephas maximus) in Thailand”. European Journal of Wildlife Research 53 (4): 298–303. doi:10.1007/s10344-007-0099-x. Retrieved 2008-04-14.
Futuyma, Douglas J; Mayer, Gregory C (1980). “Non-Allopatric Speciation in Animals”. Systematic Zoology 29 (3): 254. doi:10.2307/2412661. JSTOR 2412661.
Gavrilets, Sergey; Li, Hai; Vose, Michael (1998). “Rapid parapatric speciation on holey adaptivelandscapes” (PDF). Proc Biol Sci. 265 (1405): 1483–1489. doi:10.1098/rspb.1998.0461. PMC 1689320. PMID 9744103.
Hoskin, Conrad J; Higgie, Megan; McDonald, Keith R; Moritz, Craig (2005). “Reinforcement drives rapid allopatric speciation”. Nature 437 (7063): 1353–1356. doi:10.1038/nature04004. PMID 16251964.
Lande, Russel (1980). “Genetic Variation and Phenotypic Evolution During Allopatric Speciation”. The American Naturalist 116 (4): 463–479. doi:10.1086/283642. JSTOR 2460440.
Mayr, Ernst (1970). Populations, Species, and Evolution. The Belknap Press of Harvard University Press, Cambridge, MA. ISBN 0-674-69013-3. Retrieved 12/11/2010.